|
|
| Home |
|
Prevention & Treatment of Cancers Research Projects: |
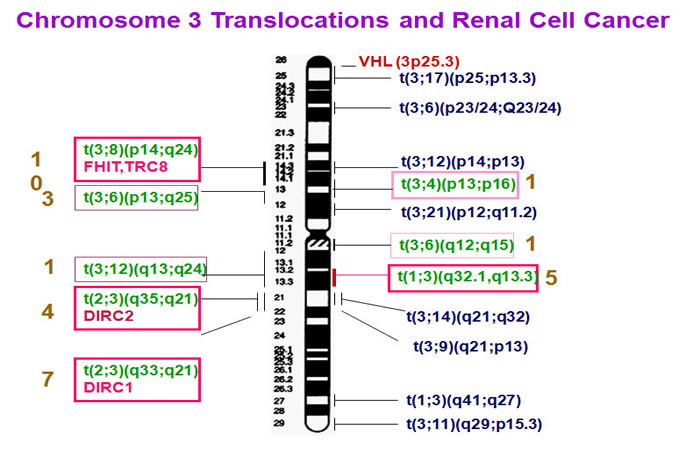
Studies of familial CCRCC have led to the identification of important tumor suppressor genes such as VHL (von Hippel-Lindau gene). Recently, position cloning also resulted in the discovery of another kidney cancer-related gene BHD. While hereditary CCRCCs are mainly attributed to (VHL) gene mutations, there are known CCRCC families and a significant proportion of sporadic CCRCCs are not linked to the VHL gene, thus pointing to the existence of other CCRCC-related genes. Since some CCRCC families are associated with balanced chromosomal translocations, the chromosomal translocation breakpoint-spanning genes are likely CCRCC-related candidate genes. The first CCRCC-associated family with a balanced chromosomal translocation t(3;8)(p14;q24) was described by Cohen et al. (1979). To date, at least eight hereditary CCRCC-related balanced chromosome translocation families have been reported (Cohen et al., 1979; Kovacs et al., 1988; Kovacs et al., 1989; Koolen et al., 1998; van Kessel et al., 1999; Podolski et al., 2001; Kanayama et al., 2001). Interestingly, translocation in all these CCRCC families is linked to chromosome 3, setting constitutional chromosome 3 translocation as a predisposing factor (vas Kessel et al., 1999; Bodmer et al., 1998; Bodmer et al., 2002b). The subsequent observation of the loss of translocation derivative chromosome 3 (der(3) chromosome) and somatic VHL mutations in a proportion of familial tumors led to the proposition of a three-step model of CCRCC tumorigenesis (Schmidt et al., 1995; Bodmer et al., 1998; Bodmer et al., 2002b): initial constitutional chromosome 3 translocation, subsequent somatic loss of the der(3) chromosome leading to the loss of a copy of VHL, and a third hit in the form of random somatic mutation in the second VHL allele. However, loss of the der(3) chromosome was observed only in a subset of the examined samples. Most of the analyzed familial tumors with loss of the der(3) did not carry VHL mutations. Furthermore, neither der(3) loss nor VHL mutations were observed in several tumor biopsies in the affected families (Eleveld et al., 2001; Bodmer et al., 2002a; Bodmer et al., 2002b). These observations suggest that the breakpoint-spanning genes in the familial RCC-associated chromosome 3 translocations are also likely implicated in RCC tumorigenesis or act synergistically in the above model in the form of genetic and/or epigenetic alternations, including gene mutation, gene disruption, and promoter CpG islands hypermethylation.
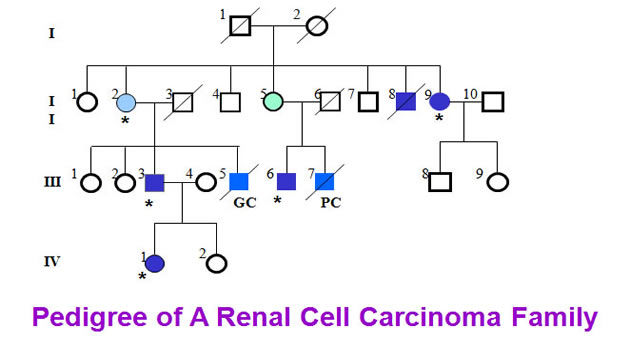
Here we collected a kidney cancer family from Japan. Five members of the family have been found to have kidney cancers. H&E staining demonstrated the patients were affected with clear cell renal cell carcinoma.
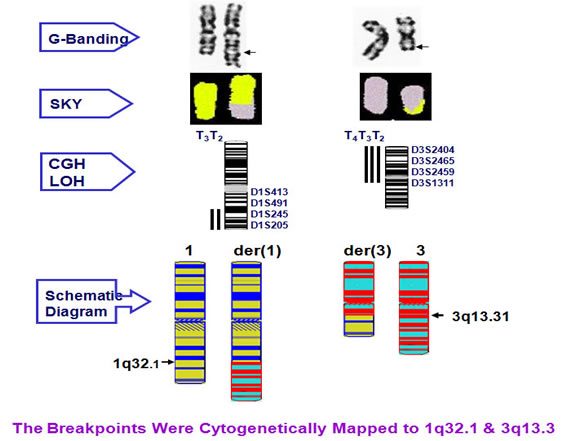
Cytogenetic analysis showed that the translocation breakpoints were located at 1q32.1 and 3q13.3 respectively.
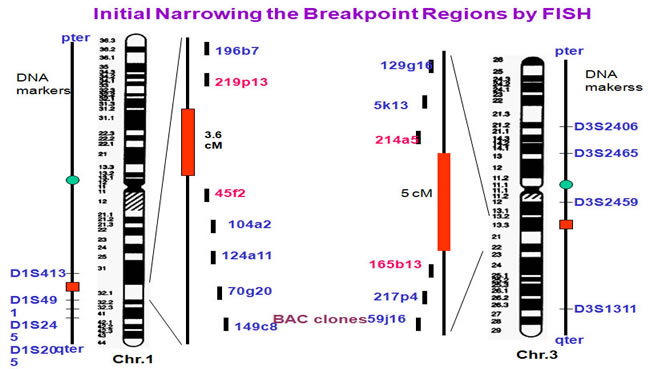
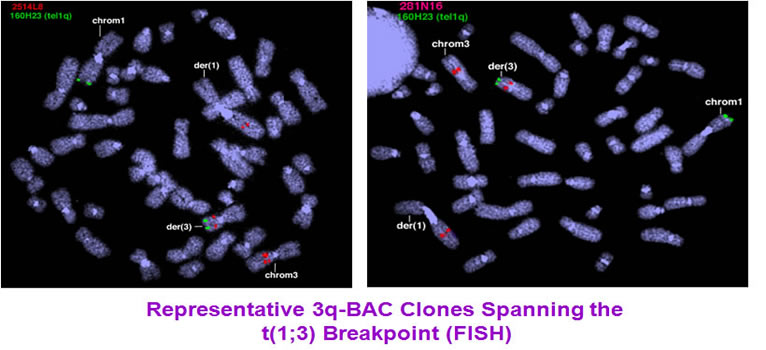
To identify the breakpoints and breakpoint-related genes, we carried out FISH experiments. The initial mapping has restricted the breakpoints to 3.6 cM region at chromosome 1 and 5 cM region at chromosome 3.
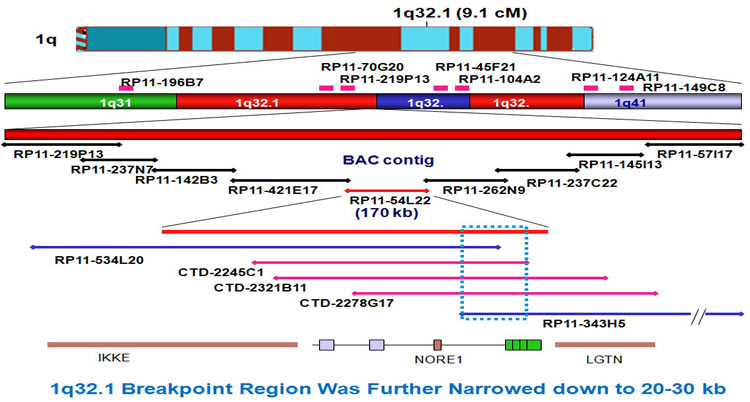
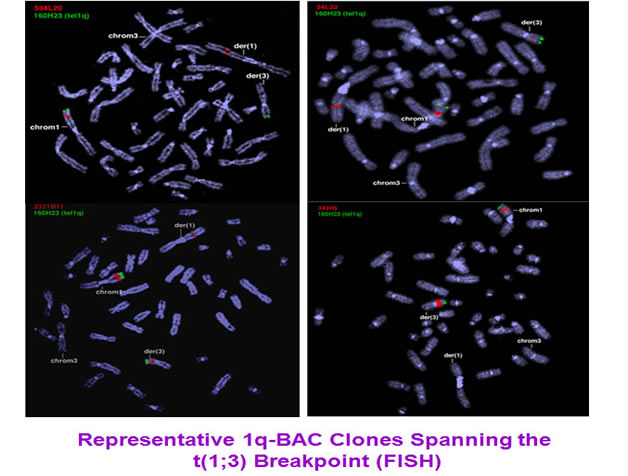
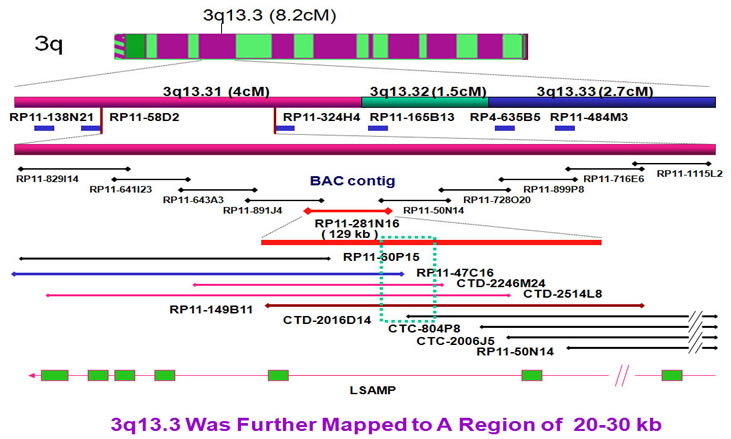
The breakpoints were further narrowed down by extended FISH contig mapping. First, we generated a 1q BAC clone contig. After FISH, the 1q region was narrowed down to a region of 20-30kb.
In same way, we set up the 3q breakpoint FISH contig to determine the 3q breakpoint region. The 3q breakpoint region was narrowed down to 20-30 kb region.
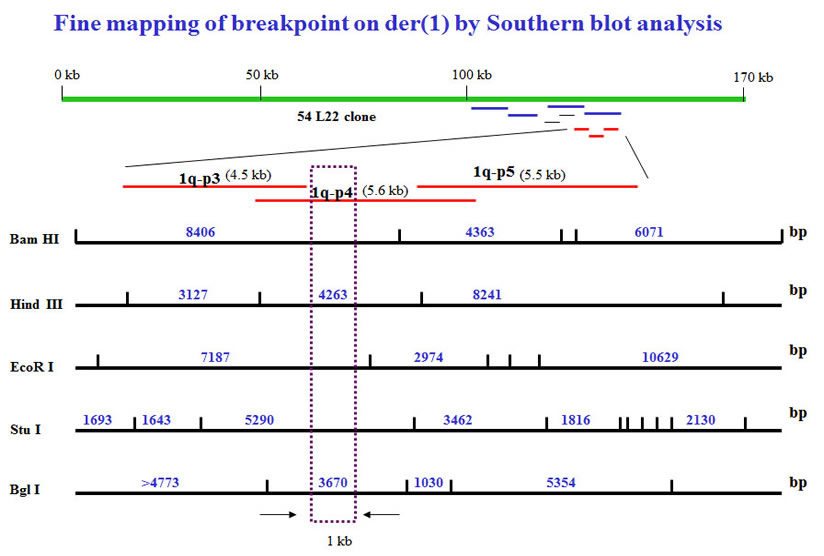
To further narrow down the breakpoints, we performed Southern blot analysis to generate restriction maps. To identify the 1q breakpoint region, we used the restriction mapping to narrow down the breakpoint to approximate 1 kb region.
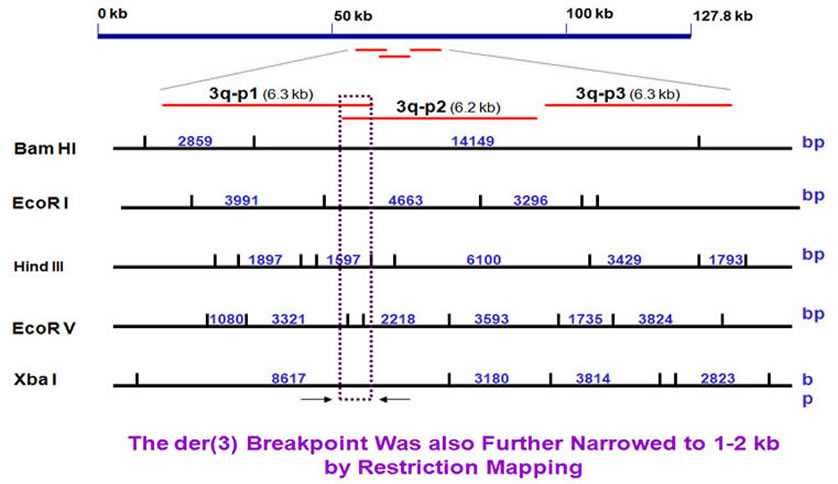
In the same way, we narrowed the 3q breakpoint region of about 1 kb.
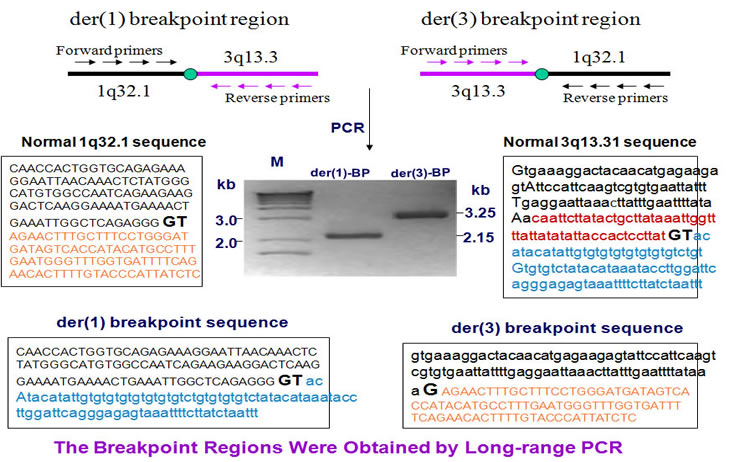
To clone the breakpoints, we designed a few pairs of PCR primers and adopted long range PCR to amplify the breakpoints.
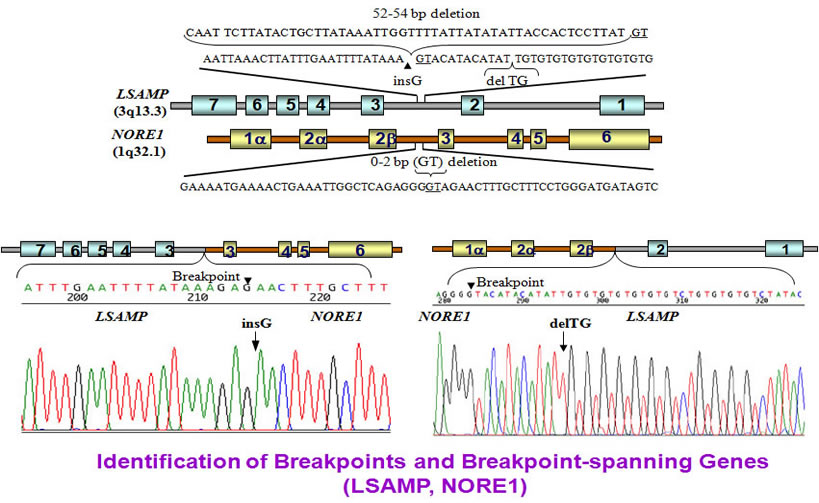
The breakpoint genes are NORE1 at chromosome 1 and LSAMP at chromosome 3. The breakpoints are located at intron2 in both genes. The LSAMP gene lost 54 bases while the NORE1 gene has a 2 bases loss.
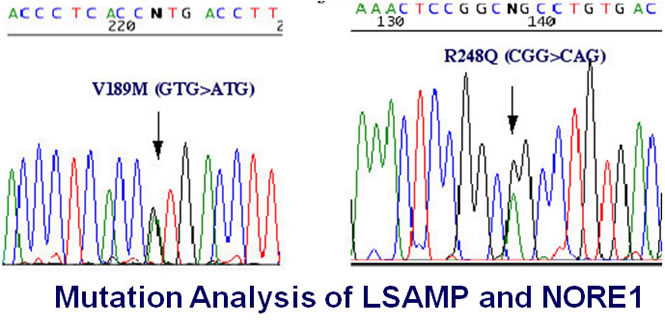
We did not found any mutations in NORE1 gene in all examined tumors and cell lines though two missense alterations were identified in LSAMP gene. It is difficult to determine these two LSAMP alterations are ture missense mutations or polymorphisms.
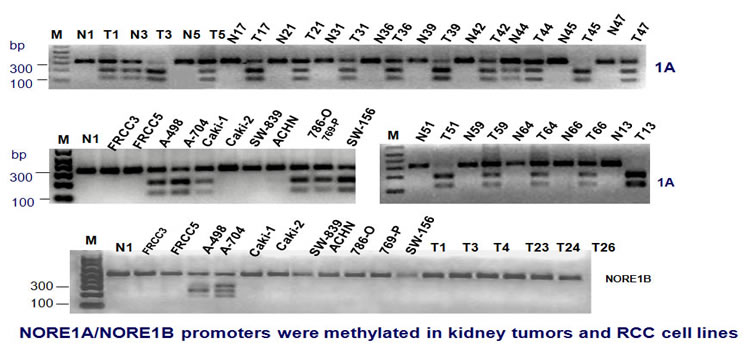
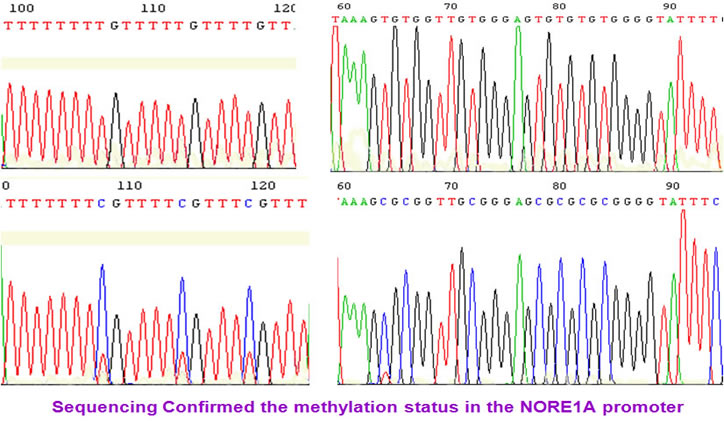
We found that NORE1A/B promoter was methylated in kidney tumors and RCC cell lines. The methylation status was confirmed by sequencing.
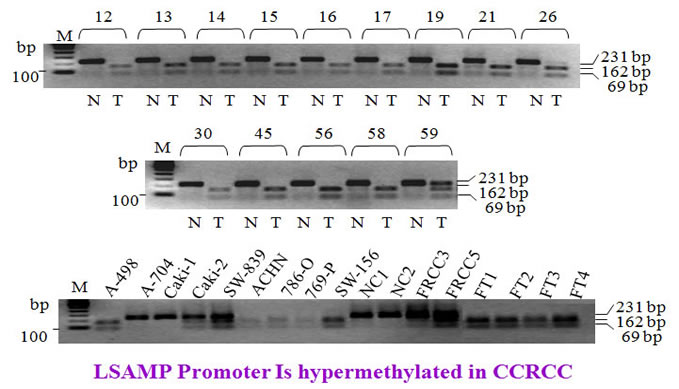
Similarly, the LSAMP promoter was methylated in RCC tumors and cell lines.
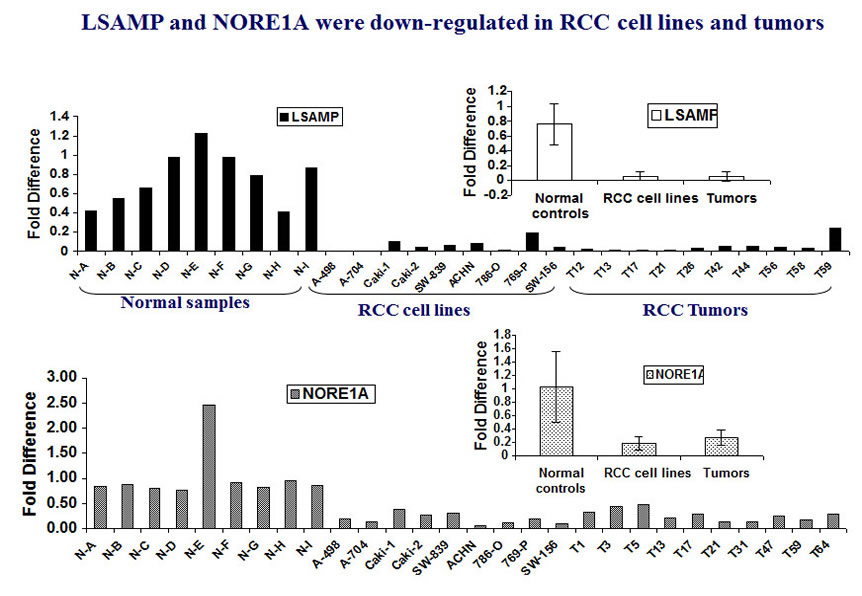
We further observed that both NORE1 and LSAMP were down-regulated in tumors and RCC cell lines. The expression level is significantly lower than that in surrounding normal tissues.
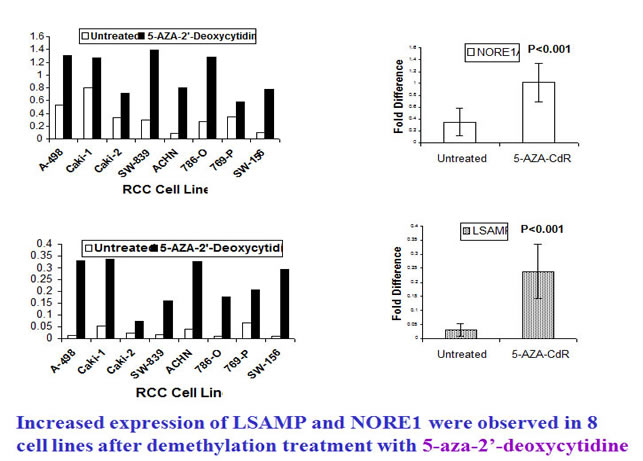
To examine whether demethylation can recover the expression of NORE1 and LSAMP, we treated the affected tumor cell lines with demethylation reagent 5-aza-2’-deoxycytidine. Clearly, the treated cell lines exhibited increased expression of NORE1 and LSAMP (below).
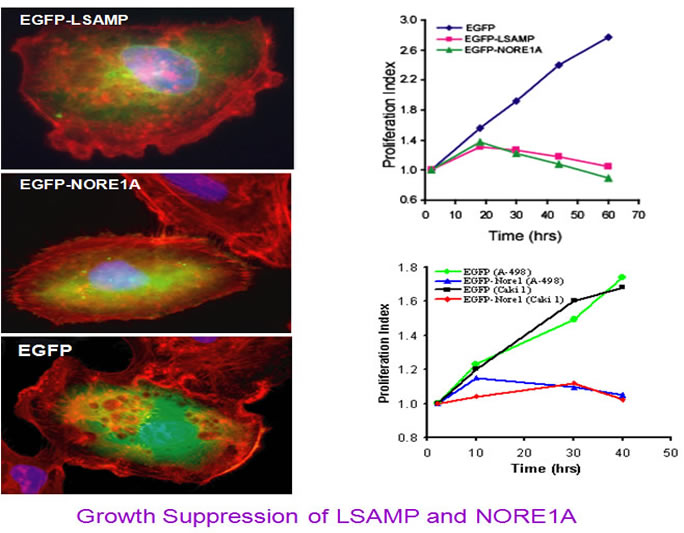
To see whether re-express the NORE1 and LSAMP proteins can inhibit tumor cell growth, we generated EGFP tagged NORE1A and LSAMP expressing plasmids and transfected into RCC cell lines. The results below showed that ectopic expression of these two proteins can effcetively inhibit cell growth.
These data suggest that LSAMP, and NORE1A (a homolog of 3p21-tumor suppressor gene RASSF1A), may represent new tumor suppressor candidates. While LSAMP-promoter hypermethylation may mainly contribute to the multi-step process of hereditary CCRCC tumorigenesis in this t(1;3)-associated family, inactivation or reduced expression of both LSAMP and/or NORE1A is likely involved in the occurrence of sporadic CCRCCs and other types of tumors. |
{kind=link}